
Porphyrins are tetrapyrrole macrocycles with conjugated double bonds and various groups attached to the perimeter. The porphyrins can accept two hydrogen ions to form +2 diacids or donate two protons to form – 2 dianions.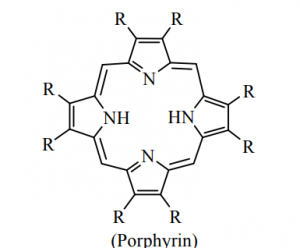 variation of substituent of facilitates the tuning of electron donating and electron withdrawing ability of the ligand.
variation of substituent of facilitates the tuning of electron donating and electron withdrawing ability of the ligand.
The porphyrin and corrin ring systems are of great biological importance. Four pyrrole units are linked by – CH = bridges as shown in figure but in corrin ring one – CH = group is less.
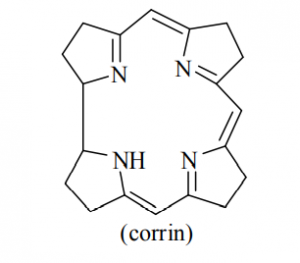
The corrin ring has 19 carbons, whereas porphyrins have 20. Also, the pyrrol like rings in corrin are fully saturated “edge-carbon” centre whereas porphyrins are highly conjugated. Because of the high number of sp3 carbon centre, corrins are more flexible than porphyrins and are not as flat. Porphyrins is aromatic in nature.
These rings are intensely colored.
Most importance of these rings in the bio-system can be illustrated as:
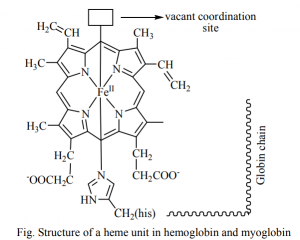
- Iron complex of the substituted porphin is Heme.
- When magnesium lies at the centre of substituted porphin ring; the resulting complex is calledchlorophyll.
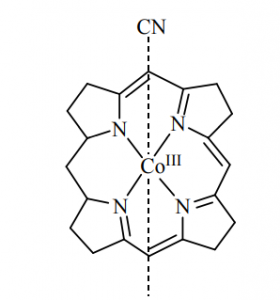
- If cobalt is the central metal atom of substituted corrin ring system, is called vitamin B12 (coblamine) Cobalt is present in +3 oxidation state in vitamin B12. Vitamin B12 is the well known naturally occurring organometallic compound. It is only vitamin known that contain metal. It may be reduced by one electron (vitamin B12 r) or two electron (vitamin B12 s) to form cobalt (II) and cobalt (I) oxidation state.
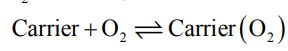
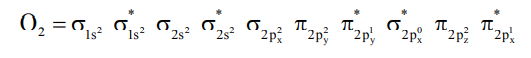

The planar porphyrin ring of heme unit of Hb and Mb, due to the presence of conjugated double bond in the porphyrin, stable ![]() and low lying
and low lying ![]() orbital are available and these allow the characteristic charge transfer electronic transition to give the red colour of blood.
orbital are available and these allow the characteristic charge transfer electronic transition to give the red colour of blood.
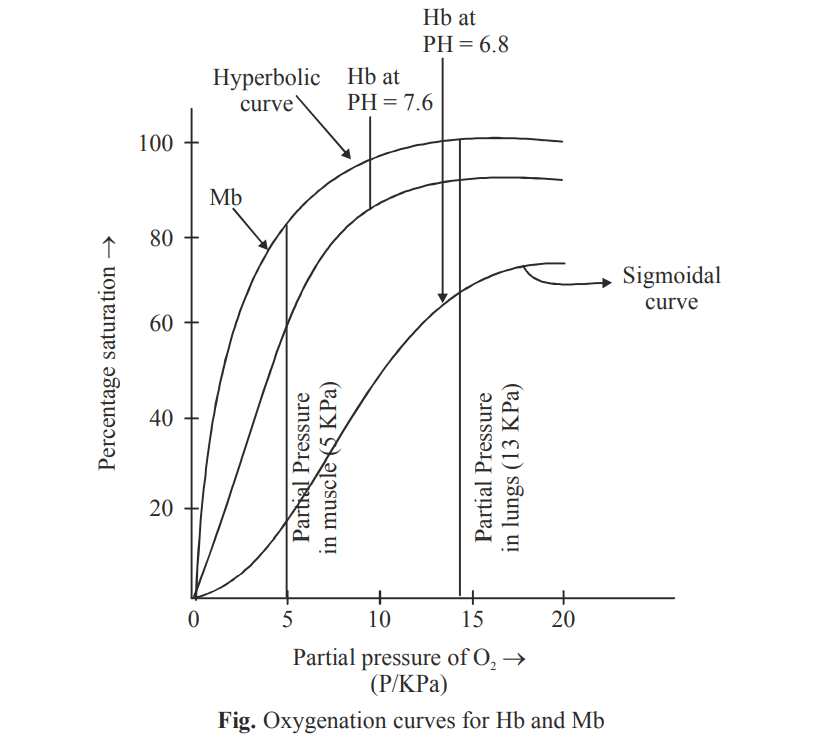

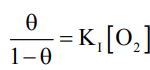
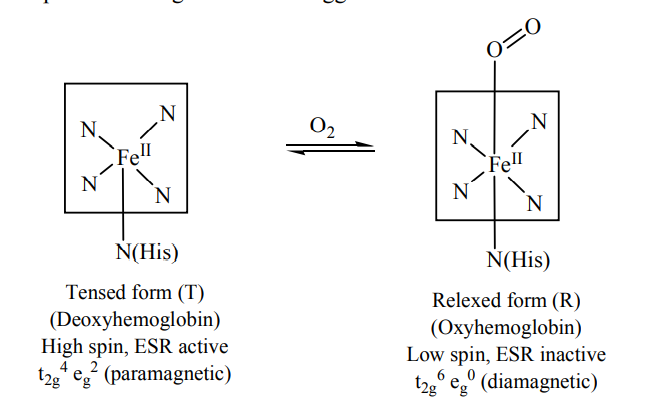
The Fe(II) – N bond length (high spin Fe(II) and sp2N) is ~218 pm. The eg electrons directly interact with the ligands and thus the removal of eg electrons in attaining the low spin stat reduces the bond length. The size of porphyrin cavity allows the sitting of the metal having the M – N bond length ~ 200-205 pm. Thus the high spin Fe(II) is deoxy Hb cannot sit in the porphyrin cavity. In fact, the size of Fe2+ increase by 28% on going from
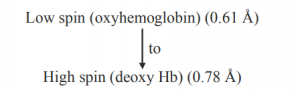 The globular protein prevents the irreversible oxidation of Fe(II) to Fe(III).
The globular protein prevents the irreversible oxidation of Fe(II) to Fe(III).
Here, it should be pointed out that there are evidences to support that the oxygenated form of Hb, iron exists as Fe(III) and O2 as O2– (superoxide) because of the, O – O stretching frequency ~ 1106 cm–1 is close to that of O– (![]() – O = 1097 cm–1). The Fe – O – O bond angle is close to 120º. In fact, Fe (II) is reversible oxidised to Fe(III) in this O2– uptake process and at the site of delivery of O2, it again attains the Fe(II) state through deoxygenation.
– O = 1097 cm–1). The Fe – O – O bond angle is close to 120º. In fact, Fe (II) is reversible oxidised to Fe(III) in this O2– uptake process and at the site of delivery of O2, it again attains the Fe(II) state through deoxygenation.
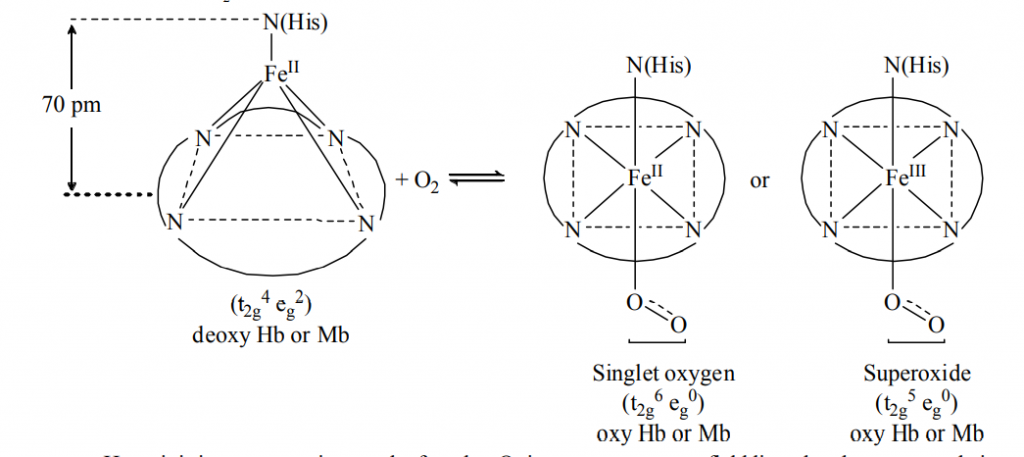
Here, it is important point out the fact that O2 is not a very strong field ligand and consequently it is not expected to carry out the desired spin pairing on oxygenation of Hb. It is believed that the heme unit is properly tuned by its substituents to make the spin pairing easier. Thus, the oxy forms of Hb and Mb, O2 remains as a singlet form. The singlet O2 is a good -acceptor and it can act as a fairly strong field ligand to induce the spin pairing in Fe(II).
Importance of glubin (protein chain):
Allosteric effect of O2, H+, CO2, Cl– and diphosphoglycerate (DPG) on O2 affinity of Hb.
Protection of Hb and Mb from irreversible oxidation by O2
Weakening the interaction of CO with heme and stabilizing the binding of O2 by histidine reversible.
![]() – donor properties of the imidazole moitey of the proximal histidine stabilise the O2 binding by the
– donor properties of the imidazole moitey of the proximal histidine stabilise the O2 binding by the ![]() – acceptance property of O2.
– acceptance property of O2.
Protonation and deprotonation sites in globin protein chain are important in maintaining the biological pH and CO2 transport.
Prevent the irreversible µ – oxo complex formation.
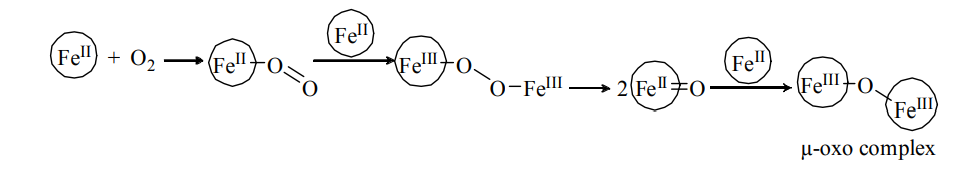
Poisoning towards hemoglobin and myoglobin:
Different ![]() -acid ligands like CO, NO, PF3 etc. which are electrically neutral and not much bulky can competitively replace O2 from the sixth octahedral site of Hb and Mb. Consequently, the O2 transport mechanism gets arrested and toxicity arieses. CN– may also bind the site, but the heme pocket surrounded by the hydrophobic environment does not welcome CN– much. The
-acid ligands like CO, NO, PF3 etc. which are electrically neutral and not much bulky can competitively replace O2 from the sixth octahedral site of Hb and Mb. Consequently, the O2 transport mechanism gets arrested and toxicity arieses. CN– may also bind the site, but the heme pocket surrounded by the hydrophobic environment does not welcome CN– much. The ![]() – bonding ligands like NH3 and amines may also block the oxygenation site.
– bonding ligands like NH3 and amines may also block the oxygenation site.
The CN– actually blocks the cytochrome C oxidase involved in the respiratory chain. To remove the bound CN– from cytochrome C oxidase, some methe-Hb (oxy-hemoglobin) are to be generated either by inhalation of amylnitrite vapour or by injection of NaNO2 solution. Met-Hb bearing Fe can bind CN– more strongly than the cytochrome C oxidase. Consequently. CN– removed from the respiratory chain to regenerate the electron tunneling path.
")
")
")
")